Me ha gustado
este capítulo, además creo que justifica con creces mi tendencia a pensar que
los virus no son seres vivos, XD! ¡Mirad la figura 2.9 y luego me contáis! (lo
sé, esto es cuestión de gustos, ya lo hemos discutido antes, pero… comparad ese
esquema simplificado de solo el metabolismo intermediario de una célula con la
frase “el genoma de los virus suele ser compacto y está muy optimizado: por lo
general, codifica únicamente su polimerasa, las proteínas de su cápside y las
necesarias para realizar funciones esenciales en el ciclo infectivo viral que
no sean aportadas por la célula hospedadora”)
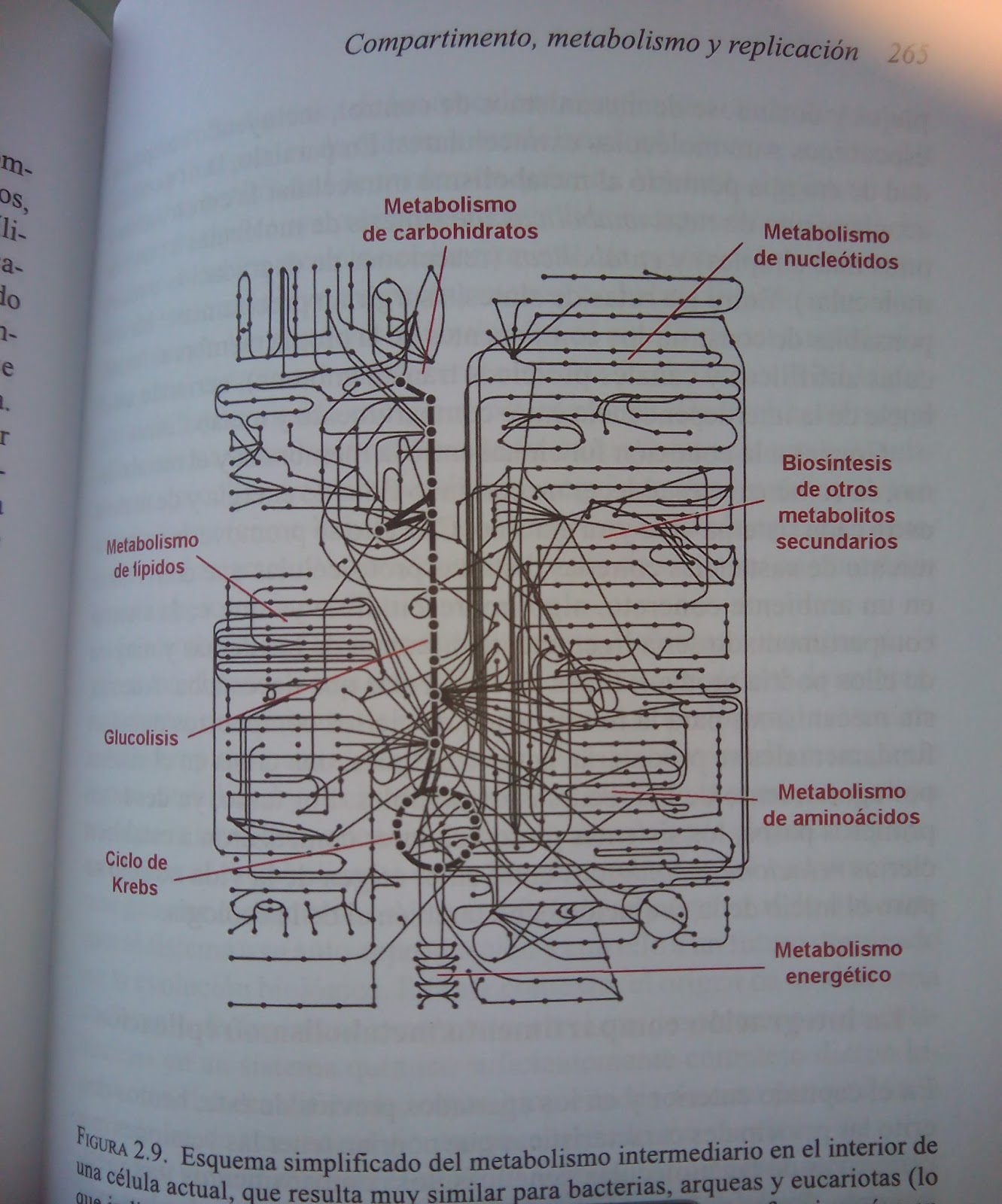
Pero que me
guste no significa que me haya resultado fácil de leer. Creo que tiene tantas
“cosicas” que muchas se me escapan, pero he pillado algunas que me han parecido
joyas (me pasó lo mismo en el de química prebiótica y en el de introducción).
¿A cuáles me refiero? A esas que, aunque son simples y lógicas, jamás las había
pensado antes.
La verdad es
que tiene que ser difícil leer este libro sin formación química y bioquímica previa
(yo solo tengo formación química y la verdad es que me cuestan algunas partes).
Por ello, en este resumen me he propuesto repasar algunos conceptos que el
autor ya introdujo en el primer capítulo (pero tal y como se pueden explicar en
los primeros cursos de la E.S .O.)
y también he echado mano del resumen que él publicó en un blog del CSIC http://blogs.20minutos.es/ciencia-para-llevar-csic/2015/12/09/el-origen-de-la-vida-cuando-la-quimica-se-convirtio-en-biologia/
Veamos como
me sale el experimento. Y si cometo errores decírmelo, que los alumnos no se
merecen que les explique mal las cosas. Si no lo veis necesario, podéis pasar
directamente al resumen del capítulo.
Resumen introductorio
Una célula
actual tiene muchas cosas, pero son partes esenciales y destacables la membrana
(formada por lípidos y proteínas), material genético (ácidos nucleicos) y el
citoplasma (medio interno líquido de la célula donde “flotan” distintas
sustancias químicas y orgánulos membranosos y no membranosos.
En las
eucariotas el material genético es ADN y está dentro del núcleo. El ADN es
copiado por ARN en el núcleo, ese ARN (mensajero) saca la información del
núcleo para la fabricación de las proteínas. Una vez fuera “trabaja” de forma
coordinada con otros ARN (el transferente, que porta los aminoácidos que darán
lugar a la proteína, y el ribosómico, que coordina y fundamenta la actuación de
todo en el ribosoma, el ribosoma es la entidad donde se sintetizan las
proteínas).
Las proteínas
son fundamentales porque muchísimas de ellas son las que controlan las
reacciones metabólicas (las reacciones en un ser vivo). Muchísimas de ellas son
enzimas (biocatalizadores) y su importancia es fundamental desde un punto de
vista termodinámico y cinético.
Una
explicación simple de cómo ocurren las reacciones químicas es la teoría de
choques: “las moléculas se mueven y, a veces, al chocar con la suficiente
fuerza y por el sitio adecuado, se rompen sus enlaces y se unen de otra manera.
La termodinámica nos habla de la estabilidad de los compuestos y de la energía
necesaria para que ocurran esas reacciones. La cinética nos habla de la
velocidad a la que ocurren esos procesos.
Siendo muy
brutos se puede decir que la vida no tiene sentido desde un punto de vista
termodinámico (por eso morimos). Entonces ¿cómo es posible? Pues porque los enzimas
(muchos de ellos proteínas) “inventan” caminos que desde un punto de vista
termodinámico son más fáciles que por los caminos sin catalizar (bajan la
energía necesaria para que ocurra la reacción) y porque la cinética de todas
las reacciones está “acoplada”, de forma que se mantienen esas moléculas inestables
asociadas a la vida el tiempo necesario para sernos útiles y que no degeneren antes
(de esta última afirmación jamás me había percatado y es una de esas ideas
simples y lógicas que nombraba en los primeros párrafos).
Toda esta
“maquinaria” metabólica la mantiene la célula gracias a un consumo de energía
(que “obtiene” del exterior) y a un consumo de sustancias químicas (que también
“toma” del exterior). Obviamente las sustancias químicas que le “sobran”
después de sus procesos son expulsadas al exterior. Otra de las ideas simples y
lógicas que me ha gustado de este capítulo es que por la membrana lipídica
pueden entrar y salir fácilmente las moléculas pequeñas, pero que las moléculas
grandes que se forman en el interior tras el metabolismo ya no salen fácilmente.
Y algo que me
ha sorprendido es que yo hago mucho hincapié cuando explico la célula en los
hidratos de carbono (en la glucosa) como fuente de energía. Les digo que en la
respiración la glucosa libera energía que es captada por las moléculas (la
fundamental el ATP) que después la cederá, por cierto muy rápidamente, a las
reacciones metabólicas de las que hemos hablado. Me ha sorprendido porque en el
capítulo pasa directamente al ATP: imagino que en los estadios iniciales de la
vida “la respiración” no es significativa (cosa obvia, porque no había O2,
aunque ya me enteraré de si esta suposición es cierta conforme avance el
libro).
Como se ha
visto en los párrafos anteriores las sustancias químicas de la vida son:
· Los
lípidos, cuya función fundamental, desde el punto de vista que aquí nos ocupa, podría
decirse que es la formación de membranas; membranas que separan el interior del
exterior y que son el soporte para que ocurran los metabolismos (lo de que las
membranas son el soporte para gran parte del metabolismo también lo he
“aprendido” en este capítulo, ¿o fue en el anterior?)
Los lípidos
más interesantes son los anfifílicos, que son los que le confieren propiedades
a membrana (doble capa lipídica). Algunos de esos lípidos anfifílicos se
pudieron formar sin problemas mediante química prebiótica (no los fosfolípidos
actuales pero si otros que pudieron hacer esas funciones de una manera menos
eficiente pero en el fondo de una manera “resultona”)
· Las
proteínas, cuya función principal, dentro de los intereses del libro, es la
catalítica.
Las proteínas
son uniones secuenciadas de aminoácidos. Los aminoácidos pudieron surgir en la
química prebiótica y algunos de ellos se pudieron unir formando péptidos de
cadena aleatoria, algunos de esos péptidos tendrían poder catalítico (pero como
vimos, los estudios actuales se inclinan a pensar en el mundo ARN)
· Los ácidos
nucleicos, cuya función principal actualmente es la genética (conservar la
información y trasmitírsela a las proteínas) y la transportadora de energía,
pero que inicialmente también podrían haber desarrollado funciones catalíticas.
Los ácidos
nucleicos son agrupaciones de nucleótidos que a su vez tienen bases
nitrogenadas. Todos esos componentes pudieron surgir de la química prebiótica y
se piensan que “evolucionaron” y generaron un mundo ARN
·Glúcidos,
cuya función principal desde el punto de vista de este libro sería la
energética (bueno, también son unos constituyentes de los nucleótidos). Aunque
esto es una apreciación mía, ya que hasta ahora no han salido mucho y está por
ver si es significativa su contribución.
Pero lo fundamental
es saber que los más simples pudieron surgir sin problemas en la química
prebiótica.
Y hasta aquí
el resumen previo al resumen del capítulo. He tenido que revisar estos
conceptos para situarme antes de leer y resumir este capítulo. Y sin más rollazo,
aquí tenéis el resumen del capítulo “compartimento,
metabolismo y replicación”
Resumen del capítulo
El capítulo
empieza diciéndonos que, aunque ya hemos hablado de la integración metabolismo-replicación
en el Mundo RNA, nos queda una tercera pata para llegar a la complejidad metabólica
que existe en la vida: el compartimento.
Además las membranas también: son "el soporte" de la captación de energía (y la posterior cesión al metabolismo) y son las responsables del trasiego de sustancias químicas del exterior al interior (y viceversa).
Y ¿por qué
triunfó la membrana doble capa lipídica?
Lo primero es que son termodinámicamente estables, no necesitan catalizadores para que ocurran (esto ya se comentó en capítulos anteriores): las micelas evolucionan a vesículas y estás pueden crecer y se parten espontáneamente cuando llegan a un tamaño crítico.
Por otro lado, dejan pasar sin problemas a moléculas orgánicas pequeñas e impiden la salida de moléculas grandes (las sintetizadas por el metabolismo en el interior de las células).
Obviamente al principio estarían formadas por lípidos mucho más simples que los actuales y con péptidos mucho más simples, pero con el paso del tiempo evolucionarían quedándose aquellos que fueran surgiendo y que le confirieran alguna propiedad que les resultara beneficiosa (es decir, de simples y poco específicas a complejas y muy específicas).
Lo primero es que son termodinámicamente estables, no necesitan catalizadores para que ocurran (esto ya se comentó en capítulos anteriores): las micelas evolucionan a vesículas y estás pueden crecer y se parten espontáneamente cuando llegan a un tamaño crítico.
Por otro lado, dejan pasar sin problemas a moléculas orgánicas pequeñas e impiden la salida de moléculas grandes (las sintetizadas por el metabolismo en el interior de las células).
Obviamente al principio estarían formadas por lípidos mucho más simples que los actuales y con péptidos mucho más simples, pero con el paso del tiempo evolucionarían quedándose aquellos que fueran surgiendo y que le confirieran alguna propiedad que les resultara beneficiosa (es decir, de simples y poco específicas a complejas y muy específicas).
Centrándonos en el metabolismo decir que, como los compuesto de la
vida no son termodinámicamente estables, tuvo que haber catalización. En el
mundo prebiótico vimos que podrían ser átomos metálicos y/o moléculas orgánicas
y/o péptidos de secuencia aleatoria. En el mundo ARN pudieron ser los
ribozimas. Pero, en la vida, han ganado los enzimas proteicos (enzimas
proteicos inicialmente poco efectivos e inespecíficos y, conforme actuara la
selección natural, muy efectivos y específicos).
Como detalle
(y aunque no lo dice, como recordatorio para los que crean en el diseño
inteligente), el autor nos recuerda que "en la fábrica del metabolismo,
las cadenas no se han ido construyendo a partir de la elegancia de un diseño ingenieril,
sino superponiendo soluciones propias del bricolaje y recurriendo a multitud de
parches que funcionaban suficientemente bien como para seguir avanzando".
Como hemos
dicho, el metabolismo es algo sumamente integrado (cinéticamente hablando y refiriéndonos
a multitud de reacciones acopladas) y por ello sobreviven los productos de la
vida (inestables, termodinámicamente hablando). Y para lograr todo esto se tuvo
que producir el acoplamiento
compartimento-metabolismo (incluyendo dentro del "concepto" compartimento
la "captación" de energía del exterior).
Siendo como
es tan importante la energía para que pueda existir el metabolismo, el autor
nos ofrece la explicación de cómo la membrana es captadora de energía.
Primero hay
que tener claro que el flujo de sustancias a través de una membrana
semipermeable libera energía y es espontaneo si ocurre favor del gradiente de
concentración (intenta igualar las concentraciones pasando sustancias).
 Pero veamos la captación de energía
para el metabolismo.
Hemos dicho que la membrana capta energía, pues bien, esa energía es usada por
la membrana para expulsar H+ y Na+ de la célula. Entonces,
como aumenta el gradiente más de lo que ya está (porque como se explicará más
adelante con el ejemplo del H+ existen más fuera que dentro siempre)
al entrar H+ se libera una energía que puede utilizarse para introducir una
sustancia que no tenga tendencia a entrar, para mover un flagelo... o para
fabricar ATP (la figura 2.8 del libro sintetiza estas ideas). Esa molécula de
ATP es la que "lleva" la energía a las reacciones metabólicas,
evolucionando a ADP que posteriormente, por un mecanismo como el que hemos
dicho, generará nuevos ATPs. Todo esto es posible por la existencia de
complejos enzimáticos (que inicialmente serían más simples) en la membrana.
Pero veamos la captación de energía
para el metabolismo.
Hemos dicho que la membrana capta energía, pues bien, esa energía es usada por
la membrana para expulsar H+ y Na+ de la célula. Entonces,
como aumenta el gradiente más de lo que ya está (porque como se explicará más
adelante con el ejemplo del H+ existen más fuera que dentro siempre)
al entrar H+ se libera una energía que puede utilizarse para introducir una
sustancia que no tenga tendencia a entrar, para mover un flagelo... o para
fabricar ATP (la figura 2.8 del libro sintetiza estas ideas). Esa molécula de
ATP es la que "lleva" la energía a las reacciones metabólicas,
evolucionando a ADP que posteriormente, por un mecanismo como el que hemos
dicho, generará nuevos ATPs. Todo esto es posible por la existencia de
complejos enzimáticos (que inicialmente serían más simples) en la membrana.
El autor nos
explica que el ATP está en todos los seres vivos (células) y que posiblemente
triunfó porque su monómero soporta mejor los ultravioletas. Pero también nos
indica que se han propuesto otras moléculas más simples que pudieron actuar de
igual forma inicialmente antes que el ATP, uno de ellos habla de una molécula
muy simple (anión pirofosfato) y otro asociado al grupo funcional -S-CO- (que
también está presente en todos los seres vivos actuales)
También el
autor nos dice que los H+ y Na+ son más abundantes en el
exterior que en el interior. ¿Por qué? Posiblemente hay más Na+
porque fueron expulsados y así se impidió que entrara agua en la célula para
diluir la alta concentración de sustancias químicas existente en ella (el agua
tiende a entrar por efecto osmótico, fenómeno a favor del gradiente). Si no
existiese ese fenómeno las proto-células hubieran reventado. Además,
coordinadamente con la salida de Na+, entrarían K+ porque
habría un desequilibrio eléctrico (esto también justificaría porque hay más K+
en el interior celular). Es decir, que todo está interrelacionado.
Y una vez
deja claro que el metabolismo complejo está integrado con la membrana, habla de
la integración de las tres cosas y los posibles modelos. Dice que los modelos
tienen que cumplir compartimentos separados, metabolismos primitivos y
moléculas genéticas (probablemente la primera fuera ARN).
Nos recuerda que como vimos en el mundo ARN la integración del metabolismo primitivo y de la replicación temprana no tienen porque ser incompatibles. Añade que si estuviera compartimentado podrían haber salido proteínas que finalmente dieran como resultado versiones más modernas de aquel metabolismo primigenio. Los modelos actuales de integración siguen distintos esquemas, pero todos análogos a los de estos tres ejemplos:
· Compartimento -> C + Metabolismo -> C + M + Replicación
· Compartimento -> C + Replicación -> C + R + Metabolismo
· Replicación -> R + Compartimento -> R + C + Metabolismo
Nos recuerda que como vimos en el mundo ARN la integración del metabolismo primitivo y de la replicación temprana no tienen porque ser incompatibles. Añade que si estuviera compartimentado podrían haber salido proteínas que finalmente dieran como resultado versiones más modernas de aquel metabolismo primigenio. Los modelos actuales de integración siguen distintos esquemas, pero todos análogos a los de estos tres ejemplos:
· Compartimento -> C + Metabolismo -> C + M + Replicación
· Compartimento -> C + Replicación -> C + R + Metabolismo
· Replicación -> R + Compartimento -> R + C + Metabolismo
 El esquema
que en la actualidad más consenso tiene sale en la figura 2.10. Además el autor
deja claro que se pudieron formar otras ramas de la vida (e incluso
diversificarse) distintas de la rama LUCA. Yo como dije en capítulos anteriores
veo esto cada vez más difícil (pero yo no cuento porque son apreciaciones sin
fundamento).
El esquema
que en la actualidad más consenso tiene sale en la figura 2.10. Además el autor
deja claro que se pudieron formar otras ramas de la vida (e incluso
diversificarse) distintas de la rama LUCA. Yo como dije en capítulos anteriores
veo esto cada vez más difícil (pero yo no cuento porque son apreciaciones sin
fundamento).
Por último
hay dos subcapítulos que hablan de líneas de investigación que se apartan en
cierto sentido de lo que hemos visto antes.
El primero habla de la química de sistemas y de que intentan generar la integración de los tres subsistemas simultáneamente desde la química prebiótica. Habla de dos o tres investigaciones pero destaca aquellas que parten de H2S y de HCN (o de HCN y Fe que formaría después el H2S) porque estarían presentes en las condiciones de la Tierra bombardeada por meteoritos y porque han logrado péptidos, ácidos nucleicos y lípidos (los precursores de los tres subsistemas). Al terminar indica que aunque es sugerente de momento no se ve muy factible el camino "simultáneo".
Y finalmente analiza que gracias a la manipulación genética se están aproximando a la vida sintética y que esto puede darnos muchas pistas de cómo surgió la vida. Nos indica que la investigación más prometedora ha sido meter en vesículas autoensambladas ARN, ADN o ácidos nucleicos artificiales y que se generen proto-células que son capaces de crecer y dividirse (buscándose ahora que estás proto-células sean metabólicamente viables y se autoreproduzcan).
El primero habla de la química de sistemas y de que intentan generar la integración de los tres subsistemas simultáneamente desde la química prebiótica. Habla de dos o tres investigaciones pero destaca aquellas que parten de H2S y de HCN (o de HCN y Fe que formaría después el H2S) porque estarían presentes en las condiciones de la Tierra bombardeada por meteoritos y porque han logrado péptidos, ácidos nucleicos y lípidos (los precursores de los tres subsistemas). Al terminar indica que aunque es sugerente de momento no se ve muy factible el camino "simultáneo".
Y finalmente analiza que gracias a la manipulación genética se están aproximando a la vida sintética y que esto puede darnos muchas pistas de cómo surgió la vida. Nos indica que la investigación más prometedora ha sido meter en vesículas autoensambladas ARN, ADN o ácidos nucleicos artificiales y que se generen proto-células que son capaces de crecer y dividirse (buscándose ahora que estás proto-células sean metabólicamente viables y se autoreproduzcan).
Despedida y cierre
Pues aquí os
dejo esto. De resumen tiene poco, la verdad.
Solo me
gustaría que si he entendido algo mal, cosa bastante probable ya que no soy
bioquímico ni biólogo, me lo digáis para que lo arregle y para que me entere yo
mejor.
Saludos.



